Molekylære maskiner
Oversatt herfra

Lenge før fremveksten av moderne teknologi sammenlignet biologistudenter livets virkemåte med maskiner.(1) I de siste tiårene har denne sammenligningen blitt sterkere enn noen gang. Som en artikkel i Nature Reviews Molecular Cell Biology sier: "I dag avslører biologi viktigheten av 'molekylære maskiner' og andre høyt organiserte molekylære strukturer som utfører de komplekse fysisk-kjemiske prosessene som livet er basert på."(2) Likeledes, en artikkel i Nature Methods observerte at "[de fleste cellulære funksjoner utføres av proteinkomplekser, som fungerer som molekylære maskiner."(3)
Hva er molekylære maskiner?
En molekylær maskin, ifølge en artikkel i tidsskriftet Accounts of Chemical Research, er "en samling av deler som overfører krefter, bevegelse eller energi fra én til en annen på en forhåndsbestemt måte."(4) En artikkel fra 2004 i Annual Review of Biomedical Engineering hevdet at "disse maskinene generelt er mer effektive enn sine motparter i makroskala," og bemerket videre at "[utallige] slike maskiner finnes i naturen." (5) Faktisk rapporterte et enkelt forskningsprosjekt i 2006 oppdagelsen av over 250 nye molekylære maskiner i gjær alene! (6)

Molekylære maskiner har utgjort en sterk utfordring for de som søker å forstå dem i darwinistiske termer som produkter av en urettet prosess. I sin bok fra 1996 Darwin's Black Box: The Biochemical Challenge to Evolution forklarte biokjemikeren Michael Behe den overraskende oppdagelsen at livet er basert på maskiner:
Kort tid etter 1950 avanserte vitenskapen til et punkt hvor den kunne bestemme formene og egenskapene til noen få av molekylene som utgjør levende organismer. Sakte, møysommelig ble strukturene til flere og flere biologiske molekyler belyst, og måten de fungerer på, utledet fra utallige eksperimenter. De kumulative resultatene viser med gjennomtrengende klarhet at livet er basert på maskiner - maskiner laget av molekyler! Molekylære maskiner frakter last fra ett sted i cellen til et annet langs "motorveier" laget av andre molekyler, mens atter andre fungerer som kabler, tau og trinser for å holde cellen i form. Maskiner slår cellulære brytere på og av, som noen ganger dreper cellen, eller får den til å vokse. Solcelledrevne maskiner fanger opp energien til fotoner og lagrer den i kjemikalier. Elektriske maskiner lar strøm flyte gjennom nerver. Produksjonsmaskiner bygger andre molekylære maskiner, så vel som seg selv. Celler svømmer ved hjelp av maskiner, kopierer seg selv med maskiner, inntar mat med maskiner. Kort sagt, svært sofistikerte molekylære maskiner kontrollerer hver cellulær prosess. Dermed er detaljene i livet fint kalibrert og livets maskineri enormt komplekst.(7)
Bilde 1. Skisse fra fotosyntese
 Behe stilte så spørsmålet: "Kan alt liv passe inn i Darwins evolusjonsteori?", og svarte: "Kompleksiteten i livets grunnlag har lammet vitenskapens forsøk på å gjøre rede for det; molekylære maskiner reiser en ennå ugjennomtrengelig barriere for darwinismens universelle rekkevidde."(8)
Behe stilte så spørsmålet: "Kan alt liv passe inn i Darwins evolusjonsteori?", og svarte: "Kompleksiteten i livets grunnlag har lammet vitenskapens forsøk på å gjøre rede for det; molekylære maskiner reiser en ennå ugjennomtrengelig barriere for darwinismens universelle rekkevidde."(8)
Selv de som er uenige i Behes svar på det spørsmålet har undret seg over kompleksiteten til molekylære maskiner. I 1998 skrev tidligere president for U.S. National Academy of Sciences Bruce Alberts den innledende artikkelen til en utgave av Cell, et av verdens beste biologitidsskrifter, som feirer molekylære maskiner. Alberts berømmet "hastigheten", "eleganse", "raffinement" og "svært organisert aktivitet" til "bemerkelsesverdige" og "fantastiske" strukturer inne i cellen. Han fortsatte med å forklare hva som inspirerte slike ord:
Hele cellen kan sees på som en fabrikk som inneholder et forseggjort nettverk av sammenlåsende samlebånd, som hver er sammensatt av et sett med store proteinmaskiner. . . . Hvorfor kaller vi de store proteinsammenstillingene som ligger til grunn for cellefunksjonen for proteinmaskiner? Nettopp fordi, som maskiner oppfunnet av mennesker for å håndtere den makroskopiske verden effektivt, inneholder disse proteinsammenstillingene svært koordinerte bevegelige deler.(9)
Bilde 2. Maskin trekker vesikkel-last
På samme måte skrev Marco Piccolini i 2000 i Nature Reviews Molecular Cell Biology at "ekstraordinære biologiske maskiner realiserer drømmen til 1600-tallets vitenskapsmenn ... at 'maskiner vil til slutt bli funnet ikke bare ukjente for oss, men også utenkelige av vårt sinn." bemerker at moderne biologiske maskiner "overgår forventningene til de tidlige livsforskerne."(10)
Noen år senere viste en oversiktsartikkel i tidsskriftet Biological Chemistry vanskelighetene evolusjonsforskere har møtt når de prøver å forstå molekylære maskiner. I hovedsak må de fornekte sine vitenskapelige intuisjoner når de prøver å takle kompleksiteten i det faktum at biologiske strukturer ser ut til å være konstruert etter skjemaene til tegninger:
Molekylærmaskiner, selv om det ofte kan virke slik, er ikke laget med en blåkopi for hånden. Likevel er biokjemikere og molekylærbiologer (og mange forskere fra andre disipliner) vant til å tenke som en ingeniør, nærmere bestemt en omvendt ingeniør. Men det er ingen tegninger ... 'Ingenting i biologi gir mening bortsett fra i lys av evolusjon': vi vet at Dobzhansky (1973) må ha rett. Men sinnet vårt, til tross for at det er et produkt av å fikse seg selv, vil merkelig nok at vi skal tenke som ingeniører.(11)
 Men gir molekylære maskiner mening i lys av ikke-styrt darwinistisk evolusjon? Er det fornuftig å benekte det faktum at maskiner viser alle tegn på at de er designet? Michael Behe hevder at faktisk molekylære maskiner oppfyller selve testen som Darwin stilte for å forfalske teorien sin, og indikere intelligent design.
Men gir molekylære maskiner mening i lys av ikke-styrt darwinistisk evolusjon? Er det fornuftig å benekte det faktum at maskiner viser alle tegn på at de er designet? Michael Behe hevder at faktisk molekylære maskiner oppfyller selve testen som Darwin stilte for å forfalske teorien sin, og indikere intelligent design.
-Darwin visste at teorien hans om gradvis evolusjon ved naturlig utvalg bar en tung byrde: "Hvis det kunne påvises at det fantes et komplekst organ som umulig kunne ha blitt dannet ved tallrike, påfølgende, små modifikasjoner, ville teorien min absolutt bryte sammen."
.. Hvilken type biologisk system kunne ikke dannes ved "mange påfølgende små modifikasjoner"? Vel, for det første, et system som er irredusibelt komplekst. Med ikke-reduserbar kompleks mener jeg et enkelt system som er sammensatt av flere samvirkende deler som bidrar til den grunnleggende funksjonen, og hvor fjerning av en hvilken som helst av delene fører til at systemet effektivt slutter å fungere.(12).
Bilde 3. Darwin motbevises ved IRK-maskiner
Molekylære maskiner er svært komplekse, og i mange tilfeller begynner vi akkurat å forstå deres indre funksjoner. Som et resultat, mens vi vet at det eksisterer mange komplekse molekylære maskiner, har til dags dato bare noen få blitt studert tilstrekkelig av biologer slik at de direkte har testet for irreduserbar kompleksitet gjennom genetiske knockout-eksperimenter eller mutasjonssensitivitetstester. Det som følger er en ikke-uttømmende liste som kort beskriver 40 molekylære maskiner identifisert i den vitenskapelige litteraturen. Den første delen vil dekke molekylære maskiner som forskere har hevdet viser irreduserbar kompleksitet. Den andre delen vil diskutere molekylære maskiner som kan være irredusibelt komplekse, men som ikke har blitt studert i nok detalj av biokjemikere ennå, til å komme med et avgjørende argument.
Valgt liste over molekylære maskiner
I. MOLEKYLÆRE MASKINER SOM FORSKERNE HAR hevdet, VISER IRREDUSIBEL KOMPLEKSITET
Bilde 4. Bakterieflagellen
 1. BAKTERIEFLAGELLUM
1. BAKTERIEFLAGELLUM
Flagellen er en roterende motor i bakterier som driver en propell til å spinne, omtrent som en påhengsmotor, drevet av ionestrøm for å drive roterende bevegelse. I stand til å spinne opp til 100 000 rpm(13), kalte en artikkel i Trends in Microbiology flagellumet "en utsøkt konstruert kjemi-osmotisk nanomaskin; naturens kraftigste rotasjonsmotor, som utnytter en transmembran ion-motorkraft for å drive en filamentøs propell."(14) På grunn av dens motorlignende struktur og indre deler, skrev en molekylærbiolog i tidsskriftet Cell, "[mer] enn andre motorer , ligner flagellen en maskin, designet av et menneske."(15) Genetiske knockout-eksperimenter har vist at E. coli-flagellen er irreduserbart kompleks med hensyn til sine omtrent 35 gener.(16) Til tross for at dette er en av de best studerte molekylære maskinene, en oversiktsartikkel fra 2006 i Nature Reviews Microbiology innrømmet at "det flagellære forskningsmiljøet knapt har begynt å vurdere hvordan disse systemene har utviklet seg."(17)
Bilde 5. Tverrsnitt av cilia
 2. EUKARYOTISK CILIUM
2. EUKARYOTISK CILIUM
Ciliet er en hårlignende eller pisklignende struktur som er bygget på et system av mikrotubuli, typisk med ni ytre mikrotubulipar og to indre mikrotubuli. Mikrotubuli er forbundet med nexin-armer og en padle-lignende bevegelse settes i gang med dynein-motorer.(18) Disse maskinene utfører mange funksjoner hos eukaryoter, som å la sædceller svømme eller fjerne fremmede partikler fra halsen. Michael Behe observerer at "padle"-funksjonen til cilium vil mislykkes hvis den mangler mikrotubuli, forbindelsesarmer eller mangler tilstrekkelige dyneinmotorer, noe som gjør den irreduserbart kompleks.(19)
3. AMINOACYL-TRNA-SYNTETASER (AARS)
aaRS-enzymer er ansvarlige for å lade tRNA-er med den riktige aminosyren slik at de kan delta nøyaktig i translasjonsprosessen. I denne funksjonen er aaRS-er en "aminoacyleringsmaskin."(20) De fleste celler krever tjue forskjellige aaRS-enzymer, ett for hver aminosyre, uten hvilke transkripsjons- /translasjonsmaskineriet ikke kunne fungere ordentlig.(21) Som en artikkel i Cell Biology International sa: " Nukleotidsekvensen er også meningsløs uten et konseptuelt translativt opplegg og fysiske ''maskinvare'-evner. Ribosomer, tRNA-er, aminoacyl-tRNA-syntetaser og aminosyrer er alle maskinvarekomponenter i Shannon-meldingsmottakeren. Men instruksjonene for dette maskineriet er i seg selv kodet i DNA og utført av 'proteinarbeidere' produsert av det maskineriet. Uten maskineriet og proteinarbeiderne kan ikke budskapet mottas og forstås. Og uten genetisk instruksjon kan ikke maskineriet settes sammen."(22) Disse komponentene danner uten tvil et ikke-reduserbar komplekst system. (23)
4. BLODSTILLINGSKASKADE
Blodkoagulasjonssystemet "er et typisk eksempel på en molekylær maskin, hvor sammenstillingen av substrater, enzymer, proteinkofaktorer og kalsiumioner på en fosfolipidoverflate markant akselererer koagulasjonshastigheten."(24) Ifølge en artikkel i BioEssays, "molekylene samhandler med celleoverflaten (molekyler) og andre proteiner for å sette sammen reaksjonskomplekser som kan fungere som en molekylær maskin."(25) Michael Behe argumenterer, basert på eksperimentelle data, at blodkoagulasjonskaskaden har en irreduserbar kjerne med hensyn til komponentene etter dens initiering-stier konvergerte.(26)
5. RIBOSOMET
Ribosomet er en "RNA-maskin"(27) som "involverer mer enn 300 proteiner og RNA"(28) for å danne et kompleks der messenger-RNA oversettes til protein, og dermed spiller en avgjørende rolle i proteinsyntesen i cellen. Craig Venter, en leder innen genomikk og Human Genome Project, har kalt ribosomet "en utrolig vakker kompleks enhet" som krever et "minimum for ribosomet rundt 53 proteiner og 3 polynukleotider", noe som førte til at noen evolusjonistiske biologer frykter at det kan være irreduserbart kompleks.(29)
6. ANTISTOFFER OG DET TILPASSE IMMUNSYSTEMET
Antistoffer er 'fingrene' til det blinde immunsystemet - de lar det skille en fremmed inntrenger fra kroppen selv.(30) Men prosessene som genererer antistoffer krever en rekke molekylære maskiner.(31) Lymfocyttceller i blodet produserer antistoffer ved å blande og matche deler av spesielle gener for å produsere over 100 000 000 varianter av antistoffer.(32) Dette 'adaptive immunsystemet' lar kroppen merke og ødelegge de fleste inntrengere. Michael Behe hevder at dette systemet er irreduserbart komplekst fordi mange komponenter må være tilstede for at det skal fungere: "Et stort repertoar av antistoffer vil ikke gjøre mye nytte hvis det ikke er noe system for å drepe inntrengere. Et system for å drepe inntrengere vil ikke gjøre mye nytte hvis det ikke er noen måte å identifisere dem på. Ved hvert trinn stoppes vi ikke bare av lokale systemproblemer, men også av kravene til det integrerte systemet."(33)
II. Ekstra molekylære maskiner
 7. SPLEISEOSOMET
7. SPLEISEOSOMET
Spleiseosomet fjerner introner fra RNA-transkripter før translasjon. I følge en artikkel i Cell, "For å gi både nøyaktighet til gjenkjennelse av reaktive spleisesteder i pre-mRNA og fleksibilitet til valg av spleisesteder under alternativ spleising, viser spleiseosomet eksepsjonell komposisjons- og strukturell dynamikk som utnyttes under substratavhengig kompleks sammenstilling, katalytisk aktivering og remodellering av aktivt sted."(34) En artikkel fra 2009 i PNAS observerte at "spleiseosomet er en massiv samling av 5 RNA-er og mange proteiner"(35) - en annen artikkel foreslår "300 forskjellige proteiner og fem RNA-er, noe som gjør det blant de mest komplekse makromolekylære maskinene som er kjent."(36)
8. F0F1 ATP-SYNTASE
Ifølge cellebiolog og molekylær maskinmodeller David Goodsell, "ATP-syntase er et av underverkene i den molekylære verden."(37) Denne proteinbaserte molekylære maskinen er faktisk sammensatt av to forskjellige roterende motorer som er forbundet med en stator: Som F0 motoren drives av protoner, snur den F1-motoren. Denne kinetiske energien brukes som en generator for å syntetisere adenosintrifosfat (ATP), det primære energibærende molekylet til celler.(38)
Bilde 6. Skisse av ATP-syntase motoren
9. BAKTERIORDOPSIN
Bacteriorhodopsin "er en kompakt molekylær maskin" som bruker denne sollysenergien til å pumpe protoner over en membran.(39) Innebygd i cellemembranen består den av syv spiralformede strukturer som spenner over membranen. Den inneholder også retinal, et molekyl som endrer form etter å ha absorbert lys. Fotoner fanget opp av netthinnen tvinges gjennom de syv heliksene til utsiden av membranen.(40) Når protoner strømmer tilbake gjennom membranen, dannes ATP.
10. MYOSIN
Myosin er en molekylær motor som beveger seg langs et "spor" - i dette tilfellet aktinfilamenter - for å danne grunnlaget for muskelbevegelse eller for å transportere last i cellen. (41) Muskler bruker molekylære maskiner som myosin for å "konvertere kjemisk energi til mekanisk energi under muskelsammen-trekning."(42) Faktisk krever muskelbevegelse den "kombinerte virkningen av billioner av myosinmotorer."(43)
11. KINESIN MOTOR
På samme måte som myosin, er kinesin en proteinmaskin som binder seg til og frakter laster ved å "krype [hånd-over-hånd] langs en mikrotubuli" i cellen.(44) Kinesiner er kraftige nok til å dra store cellulære organeller gjennom cellen så vel som vesikler eller hjelpemiddel ved montering av bipolare 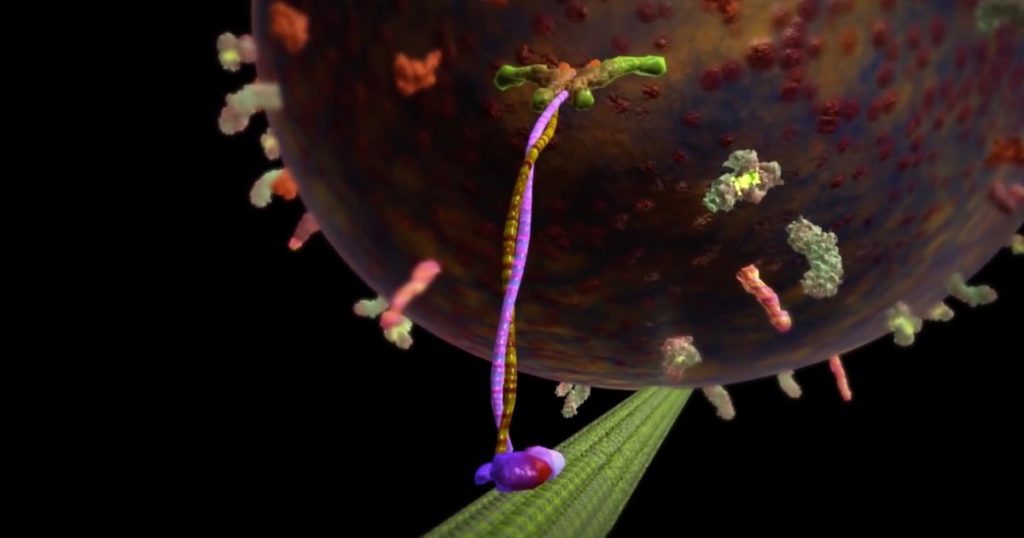 spindler, eller depolymerisering av mikrotubuli.(45)
spindler, eller depolymerisering av mikrotubuli.(45)
Bilde 7. Kinesin vandrer med sin last
12. TIM/TOM-SYSTEMER
Tim- eller Tom-systemer er selektive proteinpumpemaskiner som importerer proteiner over de indre (Tim) og ytre (Tom) membraner av mitokondriene inn i mitokondrienes indre matriks.(46)
13. KALSIUMPUMPE
Kalsiumpumpen er en "utrolig maskin med flere bevegelige deler" som overfører kalsiumioner over cellemembranen. Det er en maskin som bruker en 4-trinns syklus under pumpeprosessen.(47)
14. CYTOKROM KOKSIDASE
Cytokrom C-oksidase kvalifiserer som en molekylær maskin "siden en del av redoksfri energi omdannes til en elektrokjemisk protongradient."(48) Enzymets funksjon er å nøye kontrollere de siste trinnene i matoksidasjon ved å kombinere elektroner med oksygen og hydrogen for å danne vann, og dermed frigjøre energi. Den bruker kobber- og jernatomer for å hjelpe i denne prosessen.(49)
15. PROTEOSOM
Proteosomet er en stor molekylær maskin hvis deler må monteres nøye i en bestemt rekkefølge. For eksempel har 26S-proteosomet 33 distinkte underenheter som gjør det i stand til å utføre sin funksjon for å bryte ned og ødelegge proteiner som har blitt feilfoldet i cellen eller på annen måte merket for ødeleggelse.(50) En artikkel antydet at et bestemt eukaryotisk proteasom er kjernekomplekset i cellen" et energiavhengig proteinnedbrytningsmaskineri som tilsvarer proteinsyntesemaskineriet i sin kompleksitet."(51)
16. COHESIN
Cohesin er molekylær maskin "multisubunit protein complex"(52) og "et makromolekylært kompleks som knytter søsterkromatider sammen ved metafaseplaten under mitose."(53)
17. KONDENSIN
Condensin er en molekylær maskin som hjelper til med å kondensere og pakke kromosomer for cellereplikasjon. Det er et kompleks med fem underenheter, og er "den molekylære nøkkelmaskinen for kromosomkondensasjon." (54)
18. CLPX
ClpX er en molekylær maskin som bruker ATP til både å utfolde proteiner og deretter transportere utfoldede proteiner inn i et annet kompleks i cellen. Det flytter disse proteinene inn i ClpP-komplekset.(55)
19. IMMUNOLOGISK SYNAPSE
Den immunologiske synapsen er en molekylær maskin som fungerer som et grensesnitt for å aktivere T-celler. Når en immunologisk synapse er fullstendig dannet, aktiveres T-celler og formerer seg, noe som utløser nøkkeldelen av immunresponsen.(56)
20. GLIDEOSOME
Glideosomet er et "makromolekylært kompleks" og en "forseggjort maskin"(57) hvis funksjon er å tillate protozoer å stole på glidende motilitet over forskjellige substrater.
21. KEX2
Kex2 er en molekylær maskin som letter cellefusjon under formeringen av gjær; det virker sannsynligvis ved å degradere cellevegger.(58)
22. HSP70
Hsp70 er en av mange molekylære maskiner som fungerer som chaperoner som ikke bare hjelper andre proteiner med å oppnå en riktig funksjonell konformasjon (dvs. riktig folding), men som også hjelper dem med å bli transportert til riktig sted i cellen.(59)
23. HSP60
Hsp60 er en annen chaperone-maskin - den er skreddersydd for å gi "et lukket miljø for folding av proteiner som fullstendig beskytter dem når de folder seg".(60) Den er sammensatt av flere proteiner som danner en tønneformet struktur med en hette.(61) Når et utfoldet protein er innvendig kan den brettes skikkelig.
24. PROTEINKINASE C
Protein Kinase C er en molekylær maskin som aktiveres av visse kalsium- og diacylglycerol-signaler i cellen. Den fungerer dermed som en tolk av elektriske signaler, som en artikkel i Cell skrev: "Denne dekodingsmekanismen kan forklare hvordan cPKC-isoformer selektivt kan kontrollere forskjellige cellulære prosesser ved å stole på selektive mønstre av kalsium- og diacylglycerol-signaler."(62)
25. SECYEG PREPROTEIN TRANSLOKASJONSKANAL
SecYE-komplekset er avgjørende for driften av 'translokasjonsmaskineri' som arbeider for å flytte molekyler over membraner i cellen.(63)
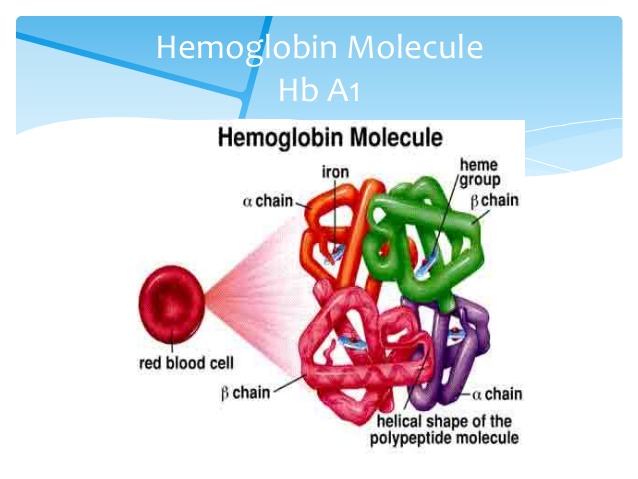 26. Hemoglobin
26. Hemoglobin
Molekylær maskinmodeller David Goodsell observerer at "Hemoglobin er en bemerkelsesverdig molekylær maskin som bruker bevegelse og små strukturelle endringer for å regulere handlingen."(64) Hemoglobin bruker jern i sin proteinstruktur for å frakte oksygen fra lungene til resten av kroppen gjennom blodet.
Bilde 8. Hemoglobin -oppbygning
27. T4 DNA-emballasjemotor
T4 DNA er en av forskjellige pakkemotorer som er "kraftige molekylære motorer" som plasserer virale genomer i kapsler kalt prokapsider.(65) Når viral genomemballasje er fullført, "frigjøres DNA-pakningsmotoren og den separat sammensatte halen festes for å produsere moden smittsom viral partikkel."(66)
28. Smc5/Smc6
Smc5/Smc6 er en kompleks maskin som er involvert i det strukturelle vedlikeholdet av kromosomer med hensyn til kohesjoner og kondensiner,(67) og arbeider for å fjerne kohesin fra skadede kromosomer før kromosomseparasjon, (68) og kan også arbeide for å reparere og løse ut DNA.(69)
29. Cytplasmatisk Dynein
Cytplasmatisk dynein er en maskin involvert i lasttransport og bevegelsescelle som fungerer som en motor med et 'kraftslag'.(70) Spesielt transporterer den kjerner i sopp og nevroner i pattedyrhjerner.(71)
30. Mitotisk spindelmaskin
Den mitotiske spindelen er en svært dynamisk selvmonterende kompleks molekylær maskin sammensatt av tubulin, motorer og andre molekyler som samles rundt kromosomene og segregerer dem til datterceller under mitose.(72)
31. DNA-polymerase
 DNA-polymerasen er en multiproteinmaskin som lager en komplementær DNA-streng fra en malstreng.(73) DNA-polymerasen er ikke bare den "sentrale komponenten i DNA-replikasjonsmaskineriet"(74), men den "spiller den sentrale rollen i livets prosesser" (75) siden det er ansvarlig for kopieringen av DNA fra generasjon til generasjon. Under polymeriseringsprosessen forblir den bundet til DNA ved hjelp av en proteinbasert glideklemme.(76) Den er ekstremt nøyaktig, og gjør mindre enn én feil per milliard baser, hjulpet av dens evne til å korrekturlese og fikse feil.(77)
DNA-polymerasen er en multiproteinmaskin som lager en komplementær DNA-streng fra en malstreng.(73) DNA-polymerasen er ikke bare den "sentrale komponenten i DNA-replikasjonsmaskineriet"(74), men den "spiller den sentrale rollen i livets prosesser" (75) siden det er ansvarlig for kopieringen av DNA fra generasjon til generasjon. Under polymeriseringsprosessen forblir den bundet til DNA ved hjelp av en proteinbasert glideklemme.(76) Den er ekstremt nøyaktig, og gjør mindre enn én feil per milliard baser, hjulpet av dens evne til å korrekturlese og fikse feil.(77)
32. RNA-polymerase
I likhet med DNA-polymerase-motstykket, er funksjonen til RNA-polymerasen å lage en messenger-RNA-streng fra en DNA-malstreng. Kalt "en enorm fabrikk med mange bevegelige deler" (78) er den en "retningsmaskin og faktisk som en molekylær motor" der den fungerer "som en dynamisk, fluktuerende, molekylær motor som er i stand til å produsere kraft og dreiemoment."(79)
Bilde 9.DNA-syntese -skjematisk
33. Kinetochore
Kinetochore er en "proteinholdig struktur som samles på sentromerisk kromatin og forbinder sentromeren til spindelmikrotubuli."(80) Kalt en "makromolekylær proteinmaskin,"(81) er den sammensatt av over 80 proteinkomponenter;(82) hjelper den med å skille kromosomer under celledeling.
34. MRX Complex
MRX-komplekset danner telomerlengde-tellemaskineri som måler integriteten til telomerer, strukturene som beskytter endene av eukaryote kromosomer. Riktig måling av telomerlengde er avgjørende for å sikre riktig cellelevetid og genomstabilitet.(83) Gjær bruker MRX-komplekset via en "proteintellings-mekanisme der høyere antall proteiner bundet av en lengre telomerrepetisjons-kanal til slutt hemmer telomeraseaktiviteten ved den aktuelle telomeren ."(84)
35. Apoptosom / Caspase
Mens mange molekylære maskiner holder en celle i live, er det til og med maskiner som er programmert til å forårsake celledød eller apoptose. Celledød må times nøye slik at cellene dør når de må erstattes. I følge David Goodsell, "caspaser er bødlene av apoptose," og de virker ved å ødelegge spesifikke proteiner i riktig rekkefølge for å "demontere cellen på en ryddig måte."(85) Caspaser kan være en del av en "dødsmaskin" kalt apoptosomet,(86) en molekylær maskin som mottar signaler som indikerer cellulær stress og deretter initierer celledød, inkludert aktivitet av caspaser.
36. Type III Sekretorisk System
Denne maskinen, ofte kalt T3SS, er en giftinjeksjonsmaskin som brukes av rovbakterier for å levere dødelige giftstoffer inn i andre celler.(87) Den er sammensatt av underenheter som er maskiner, slik som den injeksjonsbaserte nanomaskinen.(88)
37. Type II sekresjonsapparat
T2SS er en kompleks nanomaskin som translokerer proteiner over den ytre membranen til en bakterie.(89)
 38. Helicase/Topoisomerase Maskin
38. Helicase/Topoisomerase Maskin
Helikase- og topoisomerase-maskinene jobber sammen for å pakke ut eller pakke opp DNA på riktig måte før transkripsjon av DNA til mRNA eller DNA-replikasjon.(90) Topoisomerase utfører denne funksjonen ved å kutte en DNA-tråd og deretter holde på den andre mens den kuttede tråden vikles av.(91)
Bilde 10. Helicasens rolle i DNA-polymerasen
39. RNA-degradasom
RNA-degradasomet "multiproteinkompleks involvert i nedbrytningen av mRNA"(92) eller trimming av RNA til deres aktive former(93) i E. coli-bakterier. Dens store størrelse "ville lett kvalifisere [den] som en supramolekylær maskin dedikert til RNA-behandling og omsetning."(94)
40. Fotosyntetisk system
Prosessene som planter bruker for å omdanne lys til kjemisk energi en type molekylære maskiner.(95) For eksempel inneholder fotosystem 1 over tre dusin proteiner og mange klorofyll og andre molekyler som omdanner lysenergi til nyttig energi i cellen. "Antenne"-molekyler bidrar til å øke mengden lys som absorberes.(96) Mange komplekse molekyler er nødvendige for at denne banen skal fungere ordentlig.
Ovesettelse og bilder ved Asbjørn E. Lund
For referanser se nederst her